
Cada vez que alguien perfora el estípite de una palmera con un taladro para inyectarle un fitosanitario, está cometiendo un error que la palmera pagará de por vida. No porque la técnica de endoterapia sea incorrecta — es la estrategia preventiva más eficaz que existe contra el picudo rojo (Rhynchophorus ferrugineus) — sino porque la herramienta y el método no contemplan una realidad fisiológica fundamental: las palmeras no cicatrizan. Nunca. Ninguna herida en el estípite se cierra, se cubre ni se regenera. Jamás.
Este artículo explica por qué, desde la anatomía celular hasta las consecuencias fitopatológicas, y por qué esa realidad debe cambiar la forma en que se practican los tratamientos endoterápicos.
Las palmeras no son árboles
El error conceptual de fondo es tratar a las palmeras como si fueran árboles. No lo son. Son monocotiledóneas arborescentes de la familia Arecaceae — más emparentadas con los pastos, las orquídeas y los lirios que con un roble o un eucalipto — aunque su porte imponente lleve a confundirlas con árboles de forma casi inevitable.
La diferencia anatómica que lo cambia todo es el cámbium vascular. Los árboles dicotiledóneos poseen una capa cilíndrica de células meristemáticas activas entre la corteza y la madera, que genera tejido conductor nuevo de forma continua durante toda la vida del árbol. Esa capa produce xilema hacia adentro y floema hacia afuera, engrosa el tronco cada año, forma los anillos de crecimiento y, cuando el árbol es herido, genera el tejido de cierre que cubre la herida. Es la base del modelo CODIT (Compartmentalization Of Decay In Trees), descripto por Shigo y Marx (1977) y formalizado por Shigo (1984): cuando un árbol dicotiledóneo es dañado, desencadena respuestas químicas y físicas que compartimentan la lesión y construyen barreras contra la infección.
Las palmeras no tienen cámbium vascular. Sus haces vasculares (xilema y floema) están distribuidos en forma de fascículos dispersos a lo largo de todo el estípite, en lugar de concentrarse en una capa de cámbium como ocurre en los árboles dicotiledóneos. En la sección transversal del tallo se observan decenas o cientos de estos fascículos embebidos en una matriz de células parenquimáticas, con mayor concentración hacia la periferia que hacia el centro. Tomlinson (1990) en The Structural Biology of Palms -la referencia anatómica fundamental del grupo- y Broschat (2013) de la Universidad de Florida lo confirman con precisión: las heridas en el estípite que en árboles dicotiledóneos serían compartimentadas y cubiertas por tejido nuevo, en palmeras son permanentes (Broschat, 2013; DOI: 10.32473/edis-ep473-2013).
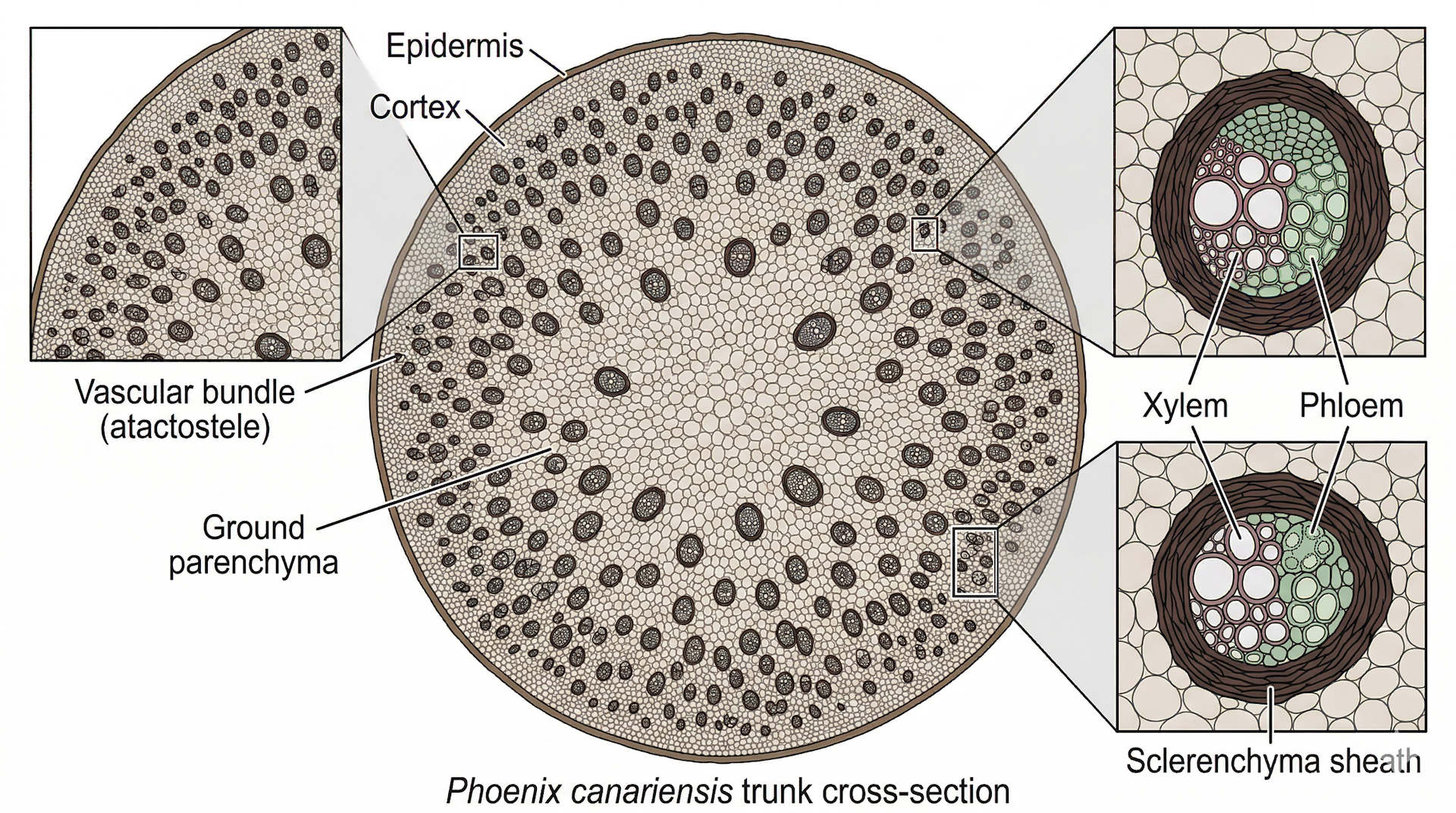
«Las palmeras no tienen meristemas laterales ni cámbium vascular. Las heridas en el estípite que en árboles dicotiledóneos serían compartimentadas y recubiertas por tejido nuevo, en palmeras son permanentes.»
Broschat, T.K. (2013). Palm Morphology and Anatomy. UF/IFAS Extension, ENH1212/EP473.
Sin cámbium no hay regeneración secundaria. Sin regeneración secundaria no hay cierre de heridas, no hay callo, no hay compartimentación. Toda herida es permanente.
La lignina: lo que hace fuertes a las palmeras también las hace vulnerables
Para entender por qué esto importa tanto, hay que entender qué es lo que da resistencia estructural a una palmera en ausencia de madera secundaria.
La respuesta es la lignina: un polímero fenólico que se deposita en las paredes celulares de las fibras esclerenquimáticas que rodean cada haz vascular. En los árboles, la lignina se concentra en la madera secundaria generada por el cámbium — tejido que muere al madurar y forma el esqueleto rígido del tronco. En las palmeras, en cambio, la lignificación ocurre dentro de los propios fascículos dispersos de xilema y floema, en las fibras que los envuelven, y continúa acumulándose durante toda la vida de la palmera, incluso en el tejido más antiguo (Rich, 1987; DOI: 10.1086/337626).
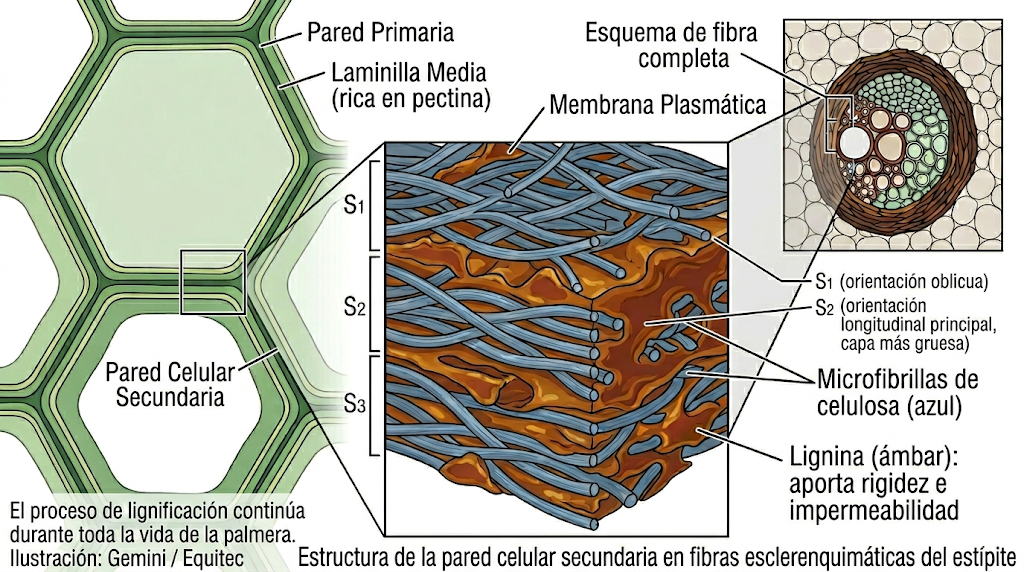
Este proceso genera además un gradiente funcional preciso: la lignificación es más intensa en la base del estípite y en su periferia — zonas de mayor estrés mecánico — y más leve hacia la punta y el centro. La base es rígida; el ápice es elástico. Por eso una palmera puede doblarse decenas de grados en un huracán sin quebrarse: no porque sea débil, sino porque su diseño distribuye la energía a lo largo de toda la columna en lugar de concentrarla en un punto de fractura. Rich (1987) documentó que la rigidez y resistencia del tejido de palmeras aumentan con la densidad seca de forma más pronunciada que en maderas convencionales, lo que explica la inusual combinación de flexibilidad y fortaleza estructural del grupo.
La paradoja es directa: la misma arquitectura de fascículos dispersos que hace a las palmeras resistentes al viento y al fuego es la que hace que sus heridas sean irreversibles. No hay cámbium que regenere. No hay tejido nuevo que cubra. Los haces vasculares dañados no se reconstituyen. El tejido necrótico no es reemplazado.
Lo que realmente ocurre cuando se perfora un estípite
Cuando se taladra el estípite de una palmera para inyectar un producto fitosanitario, ocurren varias cosas simultáneamente, ninguna de ellas reversible.
























Primero, la perforación destruye mecánicamente los haces vasculares — fascículos de xilema y floema con sus fibras lignificadas — que se encuentren en la trayectoria del taladro. Esos haces no se reconstituyen. El tejido conductor destruido es conductor perdido para siempre.
Segundo, si el sistema utiliza presión para forzar el ingreso del producto, esa presión se ejerce sobre una estructura sin cámbium y sin capacidad de regeneración. La presión mecánica puede fragmentar haces vasculares adicionales en el área circundante al punto de inserción, generando una zona de necrosis más amplia que la perforación misma. Ment et al. (2022) del Centro Volcani de Israel documentaron este daño diferencial en evaluaciones comparativas entre sistemas de endoterapia presurizada y el sistema SOSPALM.
Tercero, la herida permanece abierta de por vida. Lo que con el tiempo se observa como cavidad o hundimiento en el estípite no es necrosis activa de defensa — como sí ocurriría en un árbol dicotiledóneo mediante el modelo CODIT. Es degradación pasiva de tejido permanentemente inerte: el tejido dañado se seca, colapsa y es degradado por microorganismos porque ningún proceso regenerativo lo reemplaza.
Cuarto — y quizás lo más crítico desde el punto de vista fitopatológico — la herida abierta es una puerta de entrada permanente para patógenos vasculares. Dado que los haces de xilema y floema están distribuidos como fascículos dispersos a lo largo de toda la sección transversal, un patógeno que ingrese por esa herida tiene acceso inmediato a múltiples haces conductores sin barrera anatómica que lo contenga. Elliott (2012) de la Universidad de Florida documenta que la vía de transmisión primaria de Fusarium oxysporum f.sp. canariensis — la marchitez vascular letal de Phoenix canariensis, sin tratamiento curativo — es precisamente el uso de herramientas de poda y perforación contaminadas sobre estípites sin capacidad de cierre (Elliott, 2012; Fusarium Wilt of Canary Island Date Palm). Cada taladro sin desinfectar es un vector.
Por qué la cánula fija cambia todo
Si la herida es permanente e inevitable, porque la endoterapia requiere acceso al sistema vascular. Pero la pregunta correcta no es si perforar, sino cómo minimizar el daño y no repetirlo.
Aquí está el argumento central a favor de los sistemas de cánula fija como SOSPALM: una sola perforación bien ejecutada, con cánula que permanece colocada entre tratamiento y tratamiento, es infinitamente preferible a perforar el mismo estípite cada vez que se requiere una aplicación.

Cada perforación nueva es una herida nueva permanente. Si un programa de tratamiento preventivo contra el picudo rojo requiere aplicaciones anuales o bianuales durante años, un sistema que perfora en cada aplicación está acumulando docenas de heridas permanentes en el mismo individuo. Un sistema de cánula fija perfora una sola vez — o el mínimo posible — y reutiliza ese punto de acceso de forma controlada y sellada.
El sellado de la cánula no es un detalle menor: garantiza la oclusión del punto de perforación entre aplicaciones, reduciendo la exposición a patógenos externos y la pérdida de savia. La difusión del producto por gravedad — sin presión mecánica — permite que el principio activo se distribuya a través de los haces vasculares sin ejercer fuerzas que puedan dañar los fascículos conductores circundantes.
El contraste con la endoterapia presurizada diseñada para árboles dicotiledóneos es directo: esos sistemas fueron concebidos para estructuras con cámbium que regeneran tejido, cierran heridas y compartimentan daños. Aplicados a palmeras — organismos sin ninguna de esas capacidades — generan daño estructural adicional que la palmera jamás podrá reparar.
Tratar una palmera como si fuera un árbol no es solo un error técnico. Es un daño que la palmera cargará de por vida.
Lo que los técnicos deben saber
La fisiología de las palmeras no es un tema académico reservado a botánicos: es el fundamento de cada decisión de manejo fitosanitario. Resumida en términos prácticos:

Las palmeras no tienen cámbium vascular. Sus haces vasculares están distribuidos en forma de fascículos dispersos dentro del estípite, en lugar de concentrarse en una capa perimetral de cámbium. Eso les da resistencia mecánica extraordinaria al viento y al fuego, y les quita toda capacidad de cicatrización.
Cada herida es permanente. Cada perforación innecesaria es daño acumulado irreversible. Cada herramienta no desinfectada es un vector potencial de patógenos vasculares letales. El objetivo del tratamiento fitosanitario correcto en palmeras es proteger el ejemplar con el mínimo daño estructural posible: cánula fija, sellada, con el menor número de perforaciones necesarias, usando sistemas diseñados específicamente para la arquitectura vascular de las monocotiledóneas arborescentes.
Referencias
- Broschat, T.K. (2013). Palm Morphology and Anatomy. ENH1212/EP473. UF/IFAS Extension. https://doi.org/10.32473/edis-ep473-2013
- Elliott, M.L. (2012). Fusarium Wilt of Canary Island Date Palm. PP-215/PP139. UF/IFAS Extension. https://edis.ifas.ufl.edu/publication/PP139
- Esau, K. (1977). Anatomy of Seed Plants, 2nd ed. Wiley.
- Ment, D. et al. (2022). Evaluation of trunk injection in palm trees using SOSPALM system. Volcani Center, Israel.
- Rich, P.M. (1987). Mechanical structure of the stem of arborescent palms. Botanical Gazette, 148(1), 42–50. https://doi.org/10.1086/337626
- Shigo, A.L. (1984). Compartmentalization: A conceptual framework for understanding how trees grow and defend themselves. Annual Review of Phytopathology, 22, 189–214. https://doi.org/10.1146/annurev.py.22.090184.001201
- Shigo, A.L. & Marx, H.G. (1977). Compartmentalization of Decay in Trees. USDA Agricultural Information Bulletin 405.
- Tomlinson, P.B. (1990). The Structural Biology of Palms. Clarendon Press. https://doi.org/10.1093/oso/9780198545729.001.0001
- Tomlinson, P.B. & Huggett, B.A. (2012). Cell longevity and sustained primary growth in palm stems. American Journal of Botany, 99(12), 1891–1902. https://doi.org/10.3732/ajb.1200089

Ing. Tec. Gerardo Grinvald, Director de Equitec
Te invito a seguirnos en nuestras redes y descubrir más sobre lo que hacemos
Allí compartimos experiencias reales, testimonios y aprendizajes que reflejan nuestro trabajo día a día
Buen trabajo estimados. Ojalá qué puedan encontrar cómo eliminarlos ya que están dejando sin palmeras en muchos lados.
MUCHAS GRACIAS POR COMENTAR. CREEMOS QUE LA PREGUNTA DE COMO ELIMINAR A LA PLAGA YA ESTA RESUELTA, SIN EMBARGO HAY UN PROBLEMA MUCHO MAS GRANDE DE FALTA DE EJECUCIÓN DE UN PLAN DE MANEJO NACIONAL. GRACIAS DE NUEVO
Muy completo como siempre. Gracias muchachos por compartir sus investigaciones y trabajos en campo.
GRACIAS A TI POR SEGUIRNOS!