
El picudo rojo (Rhynchophorus ferrugineus) es conocido principalmente por los adultos que se ven en las trampas o en vuelo cerca de las palmeras. Sin embargo, es la larva el estadio responsable de la mayor parte del daño interno y de la destrucción de tejidos que conduce al colapso de la palmera: discreta, completamente oculta dentro del tronco y activa durante semanas o meses antes de que aparezca cualquier síntoma visible en el exterior. Entender qué hace esa larva, cómo progresa el daño y qué significa detectarla a tiempo es la base de cualquier decisión de manejo bien fundamentada.
La larva en el ciclo biológico
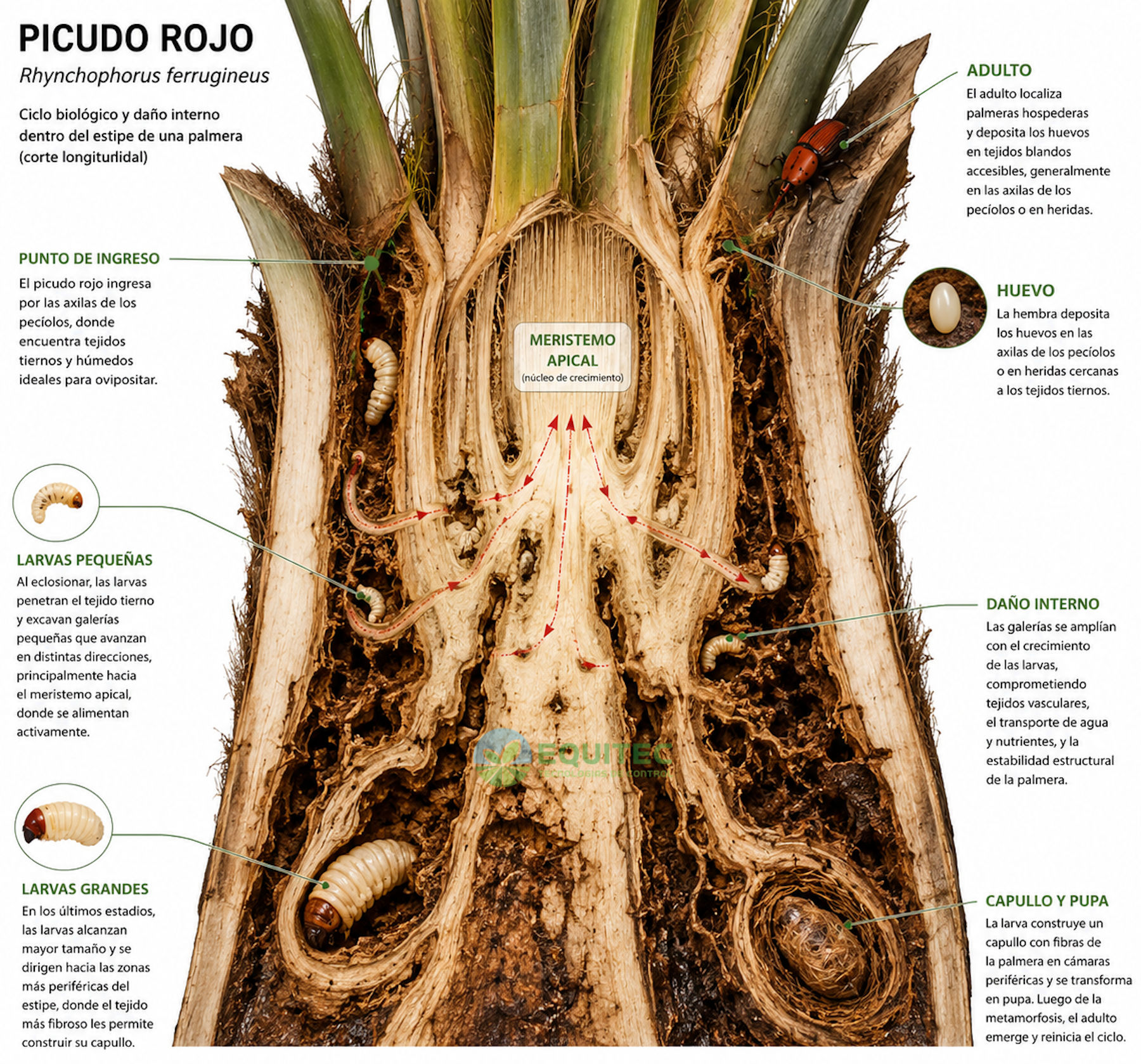
El ciclo biológico del picudo rojo tiene cuatro etapas: huevo, larva, pupa y adulto. La hembra deposita los huevos en heridas, grietas o tejidos blandos accesibles del tronco y la base foliar, utilizando el rostro para explorar y preparar sitios adecuados de oviposición. Al eclosionar, la larva penetra de inmediato en el tejido blando interior y no sale hasta completar su desarrollo. Es el estadio más largo del ciclo.
El punto de entrada y el sector de la palmera donde se establece inicialmente la infestación no son fijos. Pueden variar según la especie hospedante, la arquitectura de la palmera, la presencia de heridas previas, intervenciones de poda, el estado de estrés de la planta y la disponibilidad de tejidos blandos accesibles. Se han observado infestaciones iniciadas en bases foliares, laterales del estípite, heridas mecánicas, la zona del cuello, y en ocasiones en áreas próximas a la base del tronco cuando las condiciones lo favorecen. Esto tiene implicancias prácticas para el monitoreo: no existe un único sitio de búsqueda válido para todas las palmeras ni para todas las situaciones.
Al final del desarrollo, la larva construye un capullo compacto con fibras de la propia palmera y se transforma en pupa. El adulto que emerge puede volar activamente y alcanzar palmeras cercanas, reiniciando el ciclo.
Morfología y estadios larvarios
La larva es de color blanco cremoso, sin patas y con la cabeza de color marrón oscuro y muy endurecida. En sus últimos estadios de desarrollo llega a medir hasta 50 mm de longitud y 20 mm de diámetro, un tamaño considerable para un insecto que vive completamente dentro del tronco de una palmera. Sus mandíbulas son robustas y están adaptadas para consumir tejidos vegetales blandos.
El número de mudas que experimenta antes de convertirse en pupa no es fijo: varía según la temperatura, la especie de palmera y la dieta disponible. Dembilio y Jacas (2011) documentaron 13 estadios en Phoenix canariensis bajo clima mediterráneo. Otros estudios, realizados con dietas artificiales o en condiciones de laboratorio distintas, han reportado entre 8 y 17 estadios (Nirula, 1956; Martin y Cabello, 2006; Akutse et al., 2022). Esta variabilidad es una expresión de la plasticidad biológica de la especie frente a diferentes condiciones ambientales, no una inconsistencia de la literatura.
¿Cuánto tarda en desarrollarse?
La duración total del estadio larvario varía entre 24 y 210 días dependiendo principalmente de la temperatura (Butani, 1975; Salama et al., 2009, citados en EPPO Global Database). A temperaturas en torno a 27-28°C, el desarrollo es más rápido y la larva alcanza mayor tamaño final.
Para entender por qué la temperatura importa tanto, es útil saber que los insectos no avanzan en su desarrollo por el simple paso de los días, sino por el calor acumulado por encima de su temperatura mínima de desarrollo (estimada en aproximadamente 14-15°C para R. ferrugineus). Los científicos cuantifican ese calor útil en una unidad llamada grado-día: cada día en que la temperatura supera el mínimo aporta tantos grados-día como grados de diferencia existan entre la temperatura real y ese umbral.
Dembilio y Jacas (2011) determinaron que el picudo rojo necesita acumular 666,5 grados-día para completar su desarrollo preimaginal completo en Phoenix canariensis en clima mediterráneo. En la práctica, esto significa que en un verano con temperaturas medias de 25°C el ciclo puede completarse en unos dos meses, mientras que en otoño, con temperaturas de 18-20°C, puede llevar el triple de tiempo. En regiones con inviernos relativamente suaves, como el litoral atlántico de Uruguay, el desarrollo puede enlentecerse considerablemente sin detenerse completamente.
Aspecto en investigación: desarrollo prolongado y posibles estados de diapausa
Durante años se asumió que el enlentecimiento del desarrollo en invierno era simplemente una respuesta pasiva al frío, sin ningún mecanismo interno activo. El trabajo de Faleiro et al. (2021), publicado en Agronomy, aportó evidencia experimental que sugiere una realidad más compleja.


La diapausa larvaria es un estado de pausa fisiológica regulado activamente por el propio insecto, en el que se reduce el metabolismo, el consumo de oxígeno y la actividad alimentaria. Es cualitativamente distinta de una simple detención por frío. Los cambios fisiológicos asociados a estos estados de baja actividad metabólica podrían modificar la respuesta biológica a determinadas estrategias de control, aunque este aspecto requiere mayor investigación antes de extraer conclusiones operativas.
Faleiro et al. (2021) documentaron además que, dentro de una misma población, coexisten individuos que completan el desarrollo en tiempos esperables con otros que lo prolongan hasta aproximadamente 370 días, un fenómeno que los autores relacionan con la superdiapausa larvaria. Este hallazgo es relevante porque sugiere que parte de la variabilidad observada en la dinámica de las infestaciones podría tener base biológica en el propio insecto. Se trata, sin embargo, de un área de investigación todavía activa y sus implicancias prácticas para el manejo no están completamente establecidas.
Para más detalle sobre estos mecanismos, ver: Diapausa y superdiapausa larval en el picudo rojo.
Hospedantes y preferencias biológicas del picudo rojo
Rhynchophorus ferrugineus puede desarrollarse en numerosas especies de palmeras. La lista de hospedantes documentados incluye géneros como Phoenix, Washingtonia, Sabal, Livistona, Trachycarpus, Brahea, Chamaerops, Syagrus y otros, además de algunas plantas no paláceas como la caña de azúcar (Saccharum officinarum) (EPPO Global Database; Wakil et al., 2015). Sin embargo, la susceptibilidad no es uniforme entre especies ni entre individuos de la misma especie.
El insecto muestra diferentes niveles de preferencia dependiendo de factores como la disponibilidad y calidad del tejido alimentario y capacidad de albergarse con buenas temperaturas, el tamaño del meristemo y la estructura interna del estípite, el estado sanitario y el nivel de estrés de la planta y la presión poblacional local. La proximidad geográfica a focos de infestación activa puede también amplificar el riesgo para palmeras que en condiciones de baja presión no serían afectadas con la misma frecuencia.
Phoenix canariensis ha sido históricamente una de las especies más afectadas y con mayor documentación científica disponible, probablemente por la combinación de su gran abundancia en entornos urbanos y ornamentales, su volumen de tejido disponible y su arquitectura foliar, que puede facilitar el acceso a sitios de oviposición. Sin embargo, esto no debe interpretarse como una preferencia biológica absoluta ni como una regla generalizable: en contextos con alta presión poblacional, o cuando decrece la disponibilidad de hospedantes muy favorables, se ha observado en distintos países una expansión progresiva hacia especies consideradas inicialmente menos susceptibles. Este fenómeno podría relacionarse con la disponibilidad alimentaria y la densidad poblacional local, sin que implique necesariamente una adaptación genética nueva del insecto.
Para el manejo práctico, esto significa que ninguna especie de palmera en un entorno con presencia confirmada de R. ferrugineus debe considerarse completamente libre de riesgo, aunque la probabilidad y la frecuencia de infestación varíen significativamente entre especies y contextos.
Progresión del daño interno
Penetración inicial
Desde la eclosión, la larva perfora los tejidos blandos del interior de la palmera y comienza a alimentarse. Las galerías iniciales son angostas y de trayectoria irregular, y se van llenando de fibras masticadas mezcladas con excrementos larvarios, un material que retiene humedad y puede favorecer procesos de degradación secundaria del tejido circundante.
En palmeras jóvenes, las galerías pueden aparecer en hojas, pecíolos y la zona basal del punto de crecimiento. En palmeras adultas, las larvas se concentran inicialmente en las capas de tejido blando próximas al punto de entrada, habitualmente la base del tronco o la zona de inserción de las hojas inferiores.
Expansión y daño vascular
A medida que la larva crece y muda, sus galerías se amplían y ramifican. A medida que aumentan en tamaño y profundidad, pueden comprometer haces vasculares y otros tejidos funcionales involucrados en el transporte interno de agua y nutrientes desde las raíces hasta las hojas. A diferencia de los árboles comunes, las palmeras presentan una capacidad muy limitada para reemplazar tejidos conductivos destruidos, por lo que el daño acumulativo puede tener consecuencias permanentes. La pérdida progresiva de capacidad de transporte se expresa en clorosis, pérdida de turgencia y sequedad o marchitez de las hojas superiores.
El meristemo apical: el punto de no retorno
El meristemo apical es el único punto de crecimiento foliar de la palmera, ubicado en el centro de la corona a unos 60-70 centímetros por debajo del brote apical verde. Desde este meristemo se generan todas las hojas nuevas. En las palmeras monocotiledóneas no existe ningún tejido equivalente que pueda reemplazarlo. Cuando las larvas alcanzan y destruyen extensamente ese tejido, las posibilidades de recuperación disminuyen drásticamente.
El problema práctico es que cuando los síntomas externos hacen evidente ese daño, como la caída de las hojas centrales o el desvío del penacho, el proceso suele ser ya irreversible (Ferry y Gomez, 2002).
Masa larvaria y riesgo estructural
En infestaciones con múltiples larvas, la suma de galerías vacía progresivamente el interior meristemático. Las galerías húmedas y el tejido degradado pueden favorecer el desarrollo de microorganismos oportunistas y procesos de descomposición secundaria, que aceleran la degradación del tejido circundante y pueden asociarse con exudados marrones y olores fermentativos observados frecuentemente en palmeras severamente afectadas.
Esta pérdida de masa interna reduce la resistencia mecánica del tronco. En entornos urbanos, eso implica un riesgo de seguridad pública: palmeras con aspecto externo aceptable pueden tener el interior severamente comprometido y colapsar en condiciones de viento.
El tamaño de la larva cambia completamente el escenario
No todas las infestaciones representan el mismo riesgo ni exigen la misma respuesta. Una palmera con pocas larvas en estadios tempranos se encuentra en una situación biológica radicalmente distinta a otra con múltiples larvas grandes próximas al meristemo.
En la infestación temprana, el daño acumulado es mínimo, los haces vasculares todavía están en gran medida intactos, la carga de tejido degradado es baja y el meristemo apical no está comprometido. La palmera conserva capacidad de respuesta fisiológica, y la intervención tiene mayores probabilidades de éxito. En la infestación avanzada, la destrucción tisular es extensa, la descomposición secundaria está activa y el daño al meristemo puede ser ya irreversible o estar muy próximo a serlo.
Esta diferencia tiene consecuencias directas para el manejo. El objetivo técnico de un programa bien diseñado no es actuar sobre larvas grandes cuando el daño ya es severo, sino detectar la infestación cuando la población larvaria todavía es reducida y el tejido interno está mayoritariamente intacto. Intervenir en ese momento implica también menor carga de descomposición secundaria, mayor probabilidad de distribución adecuada de las moléculas aplicadas a través de tejidos todavía funcionales, y un costo operativo significativamente menor que el tratamiento de una palmera gravemente afectada.
Este concepto es el fundamento biológico de los programas de monitoreo sistemático: no se monitorea para constatar daño visible, sino para actuar antes de que ese daño exista.
Lo que la experiencia de campo nos enseñó sobre las larvas
La literatura científica explica cómo se desarrolla R. ferrugineus, pero la experiencia acumulada en campo aporta otra dimensión que los modelos biológicos no siempre capturan con suficiente claridad.
Una larva pequeña y una larva en estadio avanzado pertenecen al mismo insecto, pero representan escenarios completamente diferentes para la palmera. Cuando la infestación es temprana, el daño acumulado todavía es limitado, los tejidos funcionales están en gran medida intactos y la palmera conserva cierta capacidad de respuesta. La intervención en ese punto tiene un margen de acción real. Cuando la infestación está avanzada, la discusión ya no es únicamente cómo eliminar larvas: es evaluar cuánto daño estructural y fisiológico ya ocurrió. Pueden coexistir destrucción tisular extensa, descomposición secundaria activa y compromiso severo del meristemo, todo ello en una palmera que externamente todavía puede parecer aceptable.
Este contraste no es una abstracción teórica. Es una diferencia que determina qué opciones técnicas están disponibles, con qué probabilidad de resultado y a qué costo. El objetivo del manejo bien diseñado no consiste solamente en eliminar insectos: consiste en intervenir antes de que una pequeña población larvaria inicialmente invisible evolucione hacia un proceso capaz de comprometer la viabilidad completa de la palmera.
Detección precoz y toma de decisiones
El principal desafío en el manejo del picudo rojo es que el daño interno precede en semanas o meses a cualquier síntoma visible. Cuando los signos externos son claros, el meristemo suele estar ya comprometido y las posibilidades de recuperación son escasas.
Los sensores de detección por vibración sísmica que hemos estando utilizando en los últimos 4 años, permiten detectar patrones vibracionales compatibles con actividad larvaria dentro del tejido interno de la palmera. Esta información es sumamente útil para priorizar inspecciones y orientar tratamientos hacia los individuos con mayor probabilidad de infestación activa, antes de que el daño sea irreversible.Entiendo el punto. El sensor detecta actividad larval activa, pero no ve el daño previo que ya ocurrió, ni una reinfestación que arranca desde cero sobre tejido ya debilitado, ni los procesos patológicos asociados que el insecto puede introducir o facilitar.

Esta metodología de detección precoz tiene dos consecuencias prácticas: permite concentrar los recursos en las palmeras con señal positiva y puede aportar información adicional para reducir tratamientos innecesarios. Sin embargo, la ausencia de señal no debe interpretarse por sí sola como ausencia de riesgo.
Hay al menos tres razones para esa cautela. Primera: las larvas en estadios muy tempranos generan señales de menor amplitud que pueden no ser detectables; la detección es más confiable a partir de estadios intermedios. Segunda: una palmera que superó una infestación previa conserva el daño tisular y estructural acumulado, que no se reconstituye. Ese daño puede comprometer la respuesta fisiológica ante una reinfestación posterior, incluso si la nueva carga larvaria inicial es baja. Tercera: R. ferrugineus puede actuar como vector o facilitador de microorganismos patógenos, tanto transportados activamente por el insecto como oportunistas que colonizan el tejido degradado por las galerías. Las patologías asociadas a esos procesos, incluyendo pudriciones vasculares, infecciones fúngicas o bacterianas, pueden progresar de forma independiente a la actividad larvaria y no generan señal sísmica detectable.
El monitoreo instrumental es un componente valioso del manejo, pero debe combinarse con inspección visual periódica, evaluación del historial de cada palmera y criterio agronómico sobre el estado real del tejido interno.
El verdadero desafío del manejo del picudo rojo no consiste únicamente en eliminar larvas. Consiste en evitar que una pequeña población larvaria invisible se transforme en un proceso biológico y estructural capaz de comprometer una palmera completa.
Referencias bibliográficas
Nota: tanto la diapausa larvaria como el fenómeno de superdiapausa en R. ferrugineus están documentados experimentalmente por Faleiro et al. (2021), pero sus implicancias operativas concretas para los distintos métodos de control aún no han sido caracterizadas en estudios de campo independientes. La posible diapausa imaginal en adultos sugerida por Jacas et al. (2010) permanece como hipótesis preliminar sin confirmar.
Dembilio, Ó. y Jacas, J.A. (2011). Basic bio-ecological parameters of the invasive red palm weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae), in Phoenix canariensis under Mediterranean climate. Bulletin of Entomological Research, 101(2), 153-163. https://pubmed.ncbi.nlm.nih.gov/20822555/
Dembilio, Ó., Tapia, G.V., Téllez, M.M. y Jacas, J.A. (2012). Lower temperature thresholds for oviposition and egg hatching of the red palm weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae), in a Mediterranean climate. Bulletin of Entomological Research, 102(1), 97-102. https://www.cambridge.org/core/journals/bulletin-of-entomological-research/article/abs/lower-temperature-thresholds-for-oviposition-and-egg-hatching-of-the-red-palm-weevil-rhynchophorus-ferrugineus-coleoptera-curculionidae-in-a-mediterranean-climate/B850073C89DEA013E0599E5BD93A4DF5
Ferry, M. y Gomez, S. (2002). The red palm weevil in the Mediterranean area. Palms, 46(4), 172-178.
Faleiro, J.R. et al. (2021). Conservation strategy for palm groves: optimal chemical control model for red palm weevil, Rhynchophorus ferrugineus. Agronomy, 11(5), 920. https://www.mdpi.com/2073-4395/11/5/920
Nurashikin-Khairuddin, W. et al. (2022). A review of entomopathogenic nematodes as a biological control agent for red palm weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae). Insects, 13(3), 245. https://www.mdpi.com/2075-4450/13/3/245
Wakil, W., Romeno Faleiro, J. y Miller, T.A. (eds.) (2015). Sustainable Pest Management in Date Palm: Current Status and Emerging Challenges. Springer, Cham. https://link.springer.com/book/10.1007/978-3-319-24397-9
EPPO Global Database. Rhynchophorus ferrugineus (RHYCFE) — Datasheet. https://gd.eppo.int/taxon/RHYCFE/datasheet

Ing. Tec. Gerardo Grinvald, Director de Equitec
Te invito a seguirnos en nuestras redes y descubrir más sobre lo que hacemos
Allí compartimos experiencias reales, testimonios y aprendizajes que reflejan nuestro trabajo día a día
¿Nos sigues en Instagram?
En nuestro IG compartimos experiencias, errores y aprendizajes:

